我院研究团队在香菇转色阶段细胞核和线粒体转录组互作上取得新进展
近期,我院食用菌研究所良种创新与繁育团队以L808、L808-A2和L808-B为材料,从培养80 d、100 d和120 d菌丝体的细胞核和线粒体mRNA和lncRNA数据入手,探究了细胞质替换对香菇转色程度产生显著影响的原因。相关研究成果在国际学术期刊Journal of Agricultural and Food Chemistry(1区TOP,IF:5.7)上网络首发。

L808-A2和L808-B是由L808获得的核质杂种(图1A),它们的菌丝体拥有两套相同的细胞核基因组和一套不同的线粒体基因组。在80 d、100 d和120 d拥有不同转色面积的L808、L808-A2和L808-B呈现出不同的细胞核和线粒体基因和lncRNA表达谱(图1B),说明细胞质替换和培养时间都会影响香菇的转色程度。其中,L808、L808-A2和L808-B在不同培养时间上呈现的差异也是细胞质替换产生的结果。
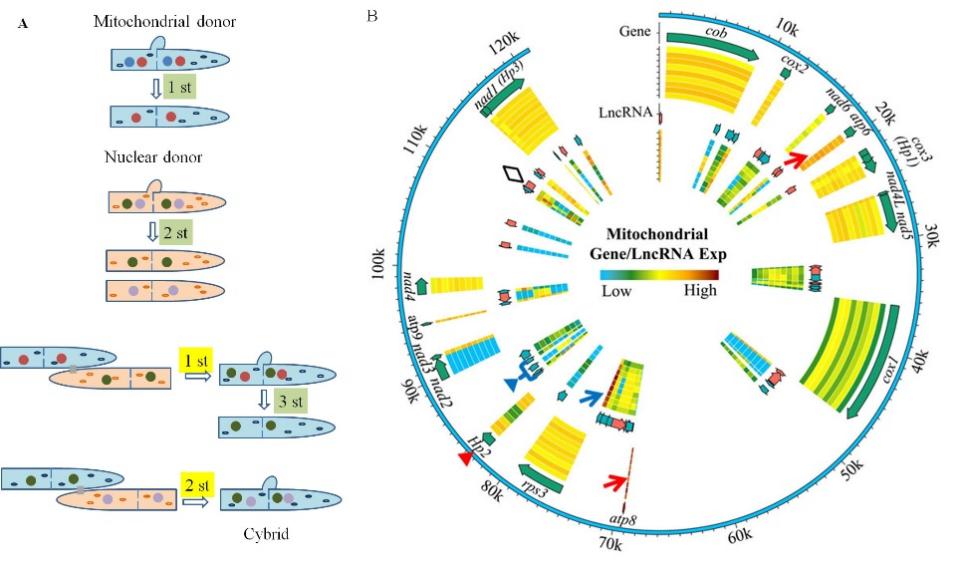
图1 香菇核质杂种获得途径(A),L808、L808-A2和L808-B在三个培养时间的线粒体基因和lncRNA表达谱(B)
通过对比分析相同培养时间(80 d、100 d或120 d)不同菌株(L808-A2 vs. L808,L808-B vs. L808,L808-B vs. L808-A2)或相同菌株(L808、L808-A2或L808-B)不同培养时间(100 d vs. 80 d,120 d vs. 100 d,120 d vs. 80 d)的细胞核和线粒体差异表达基因和lncRNAs(图2),来筛选与细胞质替换或不同培养时间显著相关的功能基因和lncRNAs。其中,在80 d、100 d和120 d时细胞质替换都显著影响了与胞外区域相关的细胞核基因,而L808、L808-A2和L808-B的不同培养时间都显著影响了与小分子代谢相关的细胞核基因(图2)。

图2 相同时间不同菌株(蓝、红、黄)和相同菌株不同培养时间(紫、绿、棕)对比分析
对三个培养时间L808、L808-A2和L808-B细胞核和线粒体的基因和lncRNAs进行共表达分析(图3A,B),筛选细胞核和线粒体在转录水平上的互作表达。结果发现与线粒体rps3基因共表达的细胞核基因数量最多,涉及多种功能(图3C)。
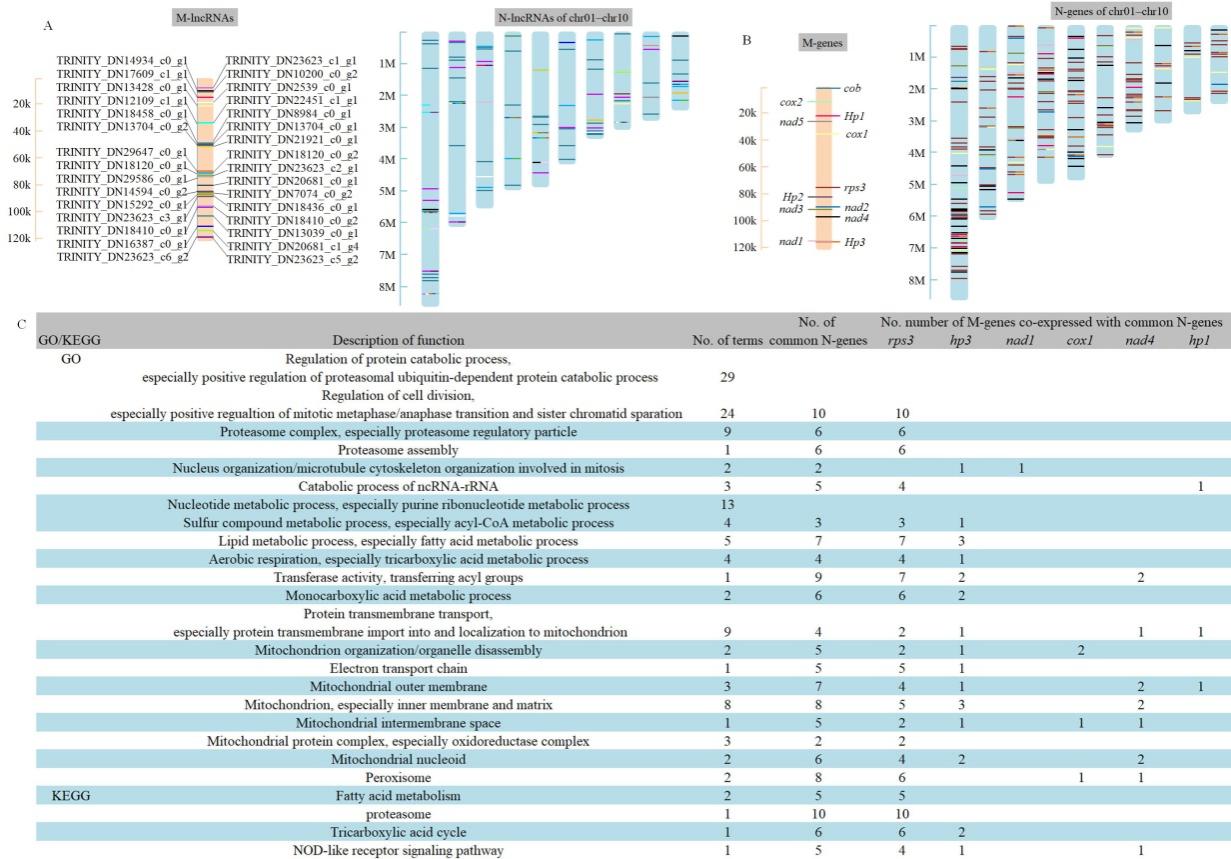
图3 细胞核和线粒体共表达的lncRNAs(A)、基因(B)和功能基因筛选(C)
为更好梳理细胞质替换对香菇转色影响的转录机制,本文通过文献整理方式绘制出了香菇转色的机制图(图4A):次级代谢和光照分别是香菇转色的基础和诱导因子;为吸收有害光,菌包表面菌丝体会积累黑色素形成褐膜,黑色素的积累涉及菌丝体胞吐作用;没有被褐膜吸收的有害光会诱导ROS产生。适量ROS可作为信号分子,最常见的是H2O2。过量的ROS会产生氧化应激,导致大分子损伤,甚至是细胞死亡。菌丝体会产生酶系或非酶系抗氧化剂以应对氧化应激,其中次级代谢产生的小分子代谢物为主要的非酶系抗氧剂,黑色素也属于非酶系抗氧化剂;NADPH介导的葡萄糖代谢为转色阶段供应能量。由此明确,细胞质替换显著影响了与香菇转色内部机制和外部褐膜形成相关的细胞核和线粒体基因,特别是一些与质膜重塑、纤维素和半纤维素胞外酶、小分子和NADPH代谢过程相关的基因(图2)。另外,结合人类rps3基因功能研究进展,明确rps3基因除了行使核糖体小亚基组装功能外,还涉及许多非核糖体的功能。本文基于L808、L808-A2和L808-B在三个培养阶段的细胞核和线粒体共表达基因富集的功能(图3C),预测了香菇线粒体rps3基因的非核糖体功能。
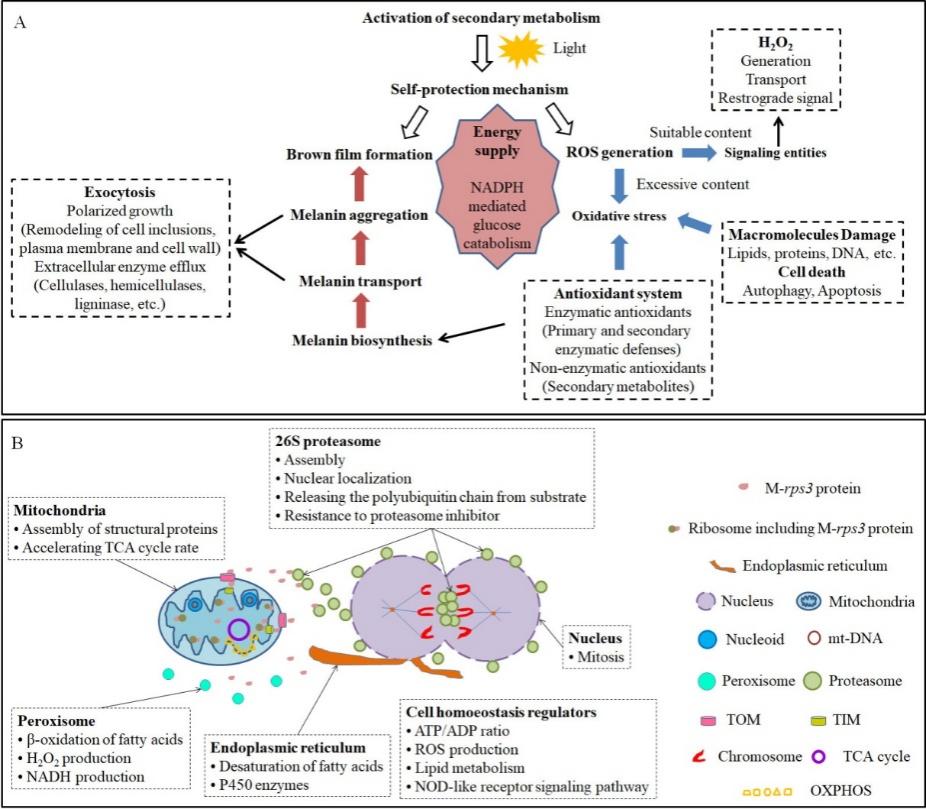
图4 香菇转色机制图(A)和线粒体rps3基因的非核糖体功能(B)
这项研究首次从细胞核和线粒体转录组互作角度对香菇转色机制进行了解析,并首次挖掘出了香菇线粒体rps3基因可能存在的非核糖体功能,使我们对香菇转色阶段、细胞核和线粒体互作都有了更深入和全面的了解。
宋晓霞副研究员和张美彦副研究员为共同第一作者,宋春艳研究员和谭琦研究员为通讯作者。该研究工作得到了国家自然科学青年基金项目的资助(32002109)。



用户登录